

Exploring Cellular Functions Using Micro Green Algae
Takako Kato-Minoura
Associate Professor, Faculty of Science and Engineering, Chuo University
Areas of Specialization: Cell Biology, Biophysics, Biochemistry, Molecular Phylogeny
The cytoskeleton, a basis of cellular function
The cytoskeleton exists within cells. Each of the cells forming our body depends on the cytoskeleton for the ability to assume certain shapes and to proliferate. Although the word "skeleton" may sound like something stiff, the cytoskeleton is actually a highly dynamic structure. Most types of cellular movement, including cell division and migration of leukocytes or cancer cells, are produced by the cytoskeleton.
Eukaryotes (all organisms except bacteria) commonly have two types of cytoskeletons: actin filaments and microtubules. The former type is composed of the protein actin (globular actin), while the latter is composed of the protein tubulin (a heterodimer of alpha- and beta-tubulin). They form three-dimensional networks within the cell (Fig. 1). Both actin and tubulin are thought to have appeared immediately after the eukaryotes first appeared on earth. Remarkably, the structure of these proteins remained almost exactly the same during the evolutionary process. (For example, the amino acid sequences of actin and tubulin are almost the same between Chlamydomonas (see below) and humans). Since cytoskeletal proteins fulfill an extremely essential function, it may have been impossible for their sequences to change even over an extremely long period of time.)
One of the most critical features of actin and tubulin is their ability to polymerize/depolymerize themselves to assemble/disassemble filaments dependent on the cellular conditions. Another valuable property is that the assembled filaments interact with force-producing proteins (motor proteins) to generate movements (using the energy supplied by ATP hydrolysis). These properties of actin and tubulin underlie the cell's morphology and motility. For example, during cell division, the chromosomes are separated into two nascent cells by the action of microtubules. Therefore, drugs that inhibit microtubule and restrict cellular proliferation are used widely as anticancer agents. At the same time, actin filaments are involved in cytokinesis, a process to partition two daughter cells after separation of chromosomes. Actin and tubulin are thus crucial proteins for fundamental cellular functions. However, many aspects remain to be studied regarding the relationship between their molecular structures and functions such as filament formation and motility.
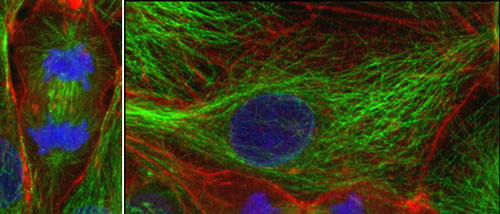
Figure 1. The cytoskeleton in the cultured animal cells
Left, a cell undergoing cell division. Actin filaments (red) and microtubules (green), and nuclei (chromosomes) specifically stained with fluorescence were observed with a confocal microscope.
Chlamydomonas, a useful organism in cytoskeleton research
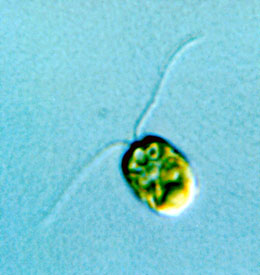
Figure 2. Chlamydomonas
Body length is about 10 μm (0.01 mm).
For our research, we use Chlamydomonas, a unicellular green alga that swims using two flagella (Fig. 2). Because it has the same chloroplasts as plants, it is also used in research on photosynthesis. Furthermore, it is a model organism for studies on cytoskeleton and cilia/flagella because its flagellar structure is common to the structure of cilia or flagella in other organisms, including humans. Many important discoveries have been made using Chlamydomonas.
The main reason why Chlamydomonas excels as an experimental model organism is that it has only one set of chromosomes in the cell (many other organisms including humans have two sets of them, one paternal and the other maternal). Since there is only one set of chromosomes, if a gene is mutated, then the effect (phenotype) will immediately appear. If there are two sets of the chromosomes, on the other hand, the phenotype may not appear (because the same gene in the remaining intact chromosome tends to suppress the effect of the mutation). Chlamydomonas is thus uniquely suitable for isolation of mutants. Many studies have been carried out using a variety of mutants in essential genes for flagellar motility or photosynthesis. If you identify a gene that causes a certain mutation and the related phenotype, then you will obtain crucial clues about the function of the protein the gene encodes.
The tubulin-gene disruptants
Utilizing the merits of Chlamydomonas, our research group has performed a wide range of research on cytoskeletons. We succeeded in obtaining a mutant of the actin gene. Analysis of this mutant led us to discover various exciting properties of actin. Additionally, we started functional studies of tubulin from several years ago. This is now our main research theme.
Despite its merit for genetic studies, Chlamydomonas has a drawback as a tool to study tubulin functions. Specifically, it has two each of the alpha- and beta-tubulin genes, and the two genes of each tubulin produce alpha-tubulin protein and beta-tubulin protein with exactly the same amino acid sequence. This means that, for tubulin genes, Chlamydomonas is in the same situation as in other organisms that have two sets of chromosomes. If a mutation is caused in a single tubulin gene, the phenotype may not appear. Moreover, even if some phenotypes do arise, we will have to examine all four tubulin genes to identify the gene in which the mutation occurred. Therefore, to avoid these problems, we first searched for cells in which one of the four tubulin genes was disrupted. We searched from about 8,000 strains in which a specific genomic fragment (cassette) was inserted at random positions and obtained the disruptants of three of the four tubulin genes (collaborative research with Kyoto University). A research group in the U.S. provided the remaining clone. Eventually, we obtained all four disruptants.
Large-scale isolation of novel tubulin mutants
We crossed these mutants to get a strain with one each of alpha- and beta-tubulin genes. Using this strain, we have now started mutagenesis on a large scale. We first screened the mutagenized cell population for mutants resistant to propyzamide, a microtubule-destabilizing drug, and obtained several beta-tubulin mutants in a highly efficient manner. Each had an amino acid substitution in beta-tubulin. On the computer-predicted three-dimensional structure of the Chlamydomonas alpha- and beta-tubulin heterodimer, all of the altered amino acids are located close to each other, near the subunit border in the dimer (Fig. 3). We further obtained mutants resistant to oryzalin, another microtubule-destabilizing drug. These mutants had amino acid substitutions in the alpha-tubulin with which oryzalin had bonded (Fig. 3). In both types of tubulin mutations, resistance to propyzamide and oryzalin is likely conferred by the structural change of tubulin produced by amino acid substitution.
In this first trial, we identified more than 15 new tubulin mutants within six months. Since only five tubulin mutants of Chlamydomonas had been reported so far, our method of obtaining tubulin-deficient mutants from a strain with single alpha- and beta-tubulin genes as a parent strain is revolutionarily efficient. We will continue to screen mutants for other conditions such as hypersensitive to microtubule-destabilizing drug or motility defects, thus collecting a wider variety of tubulin mutations. Eventually, through detailed examination of the functional abnormalities and property changes caused by the alteration of a particular amino acid, we aim to achieve a comprehensive understanding for the relationship between the structure and function of tubulin. In addition to contributing to basic biological science, this knowledge will also be essential to medicine such as cancer research.
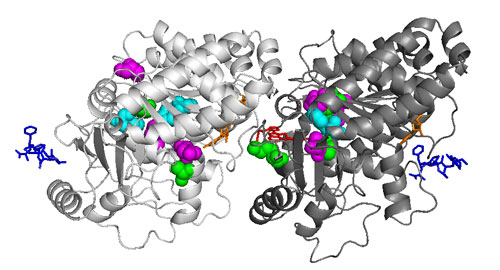
Figure 3. Structure of the Chlamydomonas tubulin heterodimer
Left, alpha-tubulin; right, beta-tubulin. The tubulin mutants isolated in our group have substitutions of different amino acids in the colored amino acids.
- Takako Kato-Minoura
Associate Professor, Faculty of Science and Engineering, Chuo University
Areas of Specialization: Cell Biology, Biophysics, Biochemistry, Molecular Phylogeny - Takako Kato-Minoura was born in Mie Prefecture in 1969. She graduated from the School of Science, Nagoya University in 1991.
In 1994, she completed the Master’s Program in the Graduate School of Science, Nagoya University.
In 1997, she finished the Doctoral Program in the same graduate school.
She holds a PhD in science from Nagoya University.
She has served as a research associate at the National Institute for Basic Biology, Okazaki National Research Institutes, technical official at the University of Tsukuba, research associate in the Graduate School of Arts and Sciences, the University of Tokyo, and assistant professor in the Faculty of Science, Gakushuin University before assuming her current position in 2008.
Her research themes include cytoskeleton protein and the morphogenesis and motility of cells in relation to cytoskeleton protein. She mainly uses cell biological methods when conducting her research. Her main written works include Basic Master Series Cell Biology (co-written, published by Ohmsha, Ltd. in 2009), Molecular Genetics: Volume 2 (published by Design Egg Inc. in 2019) and more.
- Share articles






